For years, messenger RNA was seen as a handmaiden to DNA, faithfully copying its gene-laden strands and carrying them into the cytoplasm, where they are met by other RNAs, each dedicated to the job of translating the DNA’s message. In what some are calling a paradigm shift, this story is being replaced by one with a far more tangled plot.
The shift began in 1999, when worms, flies, and other creatures were found to produce small double-stranded RNAs that tackle messenger RNA and stop it from being translated in a process called RNA interference. Researchers raced to find ways to harness these dsRNAs for the purpose of silencing genes in higher organisms. It now appears that vertebrates, including humans, produce their own bits of RNA—single-stranded jots that block or even degrade the messengers. They were first spotted in 1993 in Caenorhabditis elegans, but only recently has attention turned to this scrappy mob, the microRNAs (miRNAs).
“That such a small piece of RNA is enough to do something important—some people did not believe that,” said Changchun Xiao, an HMS research fellow in pathology.
Molecular One on OneUnlike their double-stranded RNA cousins, which may have evolved to block the activity of invading pathogens, miRNAs work in service to the cell, regulating many vital functions. Humans are thought to carry hundreds of miRNAs, each coded for by its own stretch of DNA. As much as a third of the human protein-coding genome may be affected by miRNA activity. Though they are attracting the attention of researchers in far-flung fields, these molecular regulators remain mysterious. Few have actually been caught in the act of blocking the translation of a specific messenger. Part of the problem is that they are so tiny, only 20 to 24 nucleotides long, which means they can bind to multiple targets. Indeed, microRNAs have been considered quite promiscuous, interacting with many different mRNAs.
It now appears that miRNAs, if not exactly monogamous, do engage in powerful and surprisingly intimate relationships with single mRNAs. In one of the most thorough explorations yet of a mammalian miRNA and its target, Xiao, working with colleagues including Klaus Rajewsky, the Fred S. Rosen professor of pediatrics at Children’s Hospital Boston and principal investigator at the Immune Disease Institute, found that the interaction between an miRNA and its target can have a profound effect on the cell, and it can do so in an exquisitely sensitive manner.
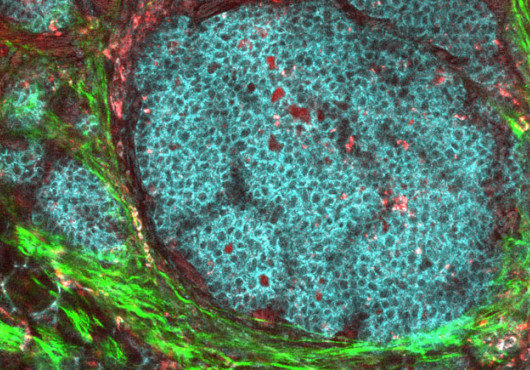
Focusing on the mouse immune system, the researchers found that altering the level of an miRNA expressed in B cells produces an equivalent though inverse change in the level of its target, in this case, the transcription factor c-Myb. These changes were reflected in the number of B cells produced by the mice. Ultimately, Xiao and colleagues were able to control the number of B1 cells produced in the mice by tweaking the amount of miRNA the cells expressed. Their findings appear in the Oct. 5 Cell.
Previous studies in C. elegans had shown that miRNAs can interact with single gene targets, essentially switching them on and off, but not that they could fine-tune gene expression so precisely. In another study appearing in the same issue of Cell, a team of researchers at the European Molecular Biology Laboratory in Germany shows that an miRNA expressed in Drosophila neurons adjusts expression of a target gene, atrophin, to an optimum level. Together, the papers “provide evidence that miRNAs join the crowd of regulatory mechanisms that tune the expression of genes to physiologically relevant levels,” writes Oliver Hobert, of Columbia University, in a commentary accompanying the Cell studies.
Part of the ProblemThe tiny scraps of RNA are already joining the ranks of putative disease agents. If they can exert such fine-scale control on critical processes inside the cell, then defects in form or levels of miRNA could wreak havoc. Indeed, miRNAs have recently been implicated in a host of diseases, including certain B cell lymphomas and metastatic breast cancer.
Rajewsky’s interest in B cells and their link to cancer dates back to long before the miRNAs were discovered. In the 1990s, he and his colleagues found that Hodgkin’s disease is caused by an errant B cell, but it was unclear what mutated genes were controlling the B cell’s aberrant behavior. Part of the problem is that normal B cell development is complex, involving many stages before cells reach maturity. “It’s a very sophisticated developmental process,” said Rajewsky. He became intrigued by the possibility that miRNAs might play a role. In 2004, he, Xiao, and colleagues joined with other labs to create a map pinpointing which miRNAs are expressed in different cells at different stages of development.
One, miR-150, emerged from the list as having an exceptionally clear expression pattern in B cells. What intrigued Xiao and Rajewsky was the way miR-150 was expressed only in resting mature cells, not in precursor cells, and not in mature cells that have been activated. To explore its role, Xiao, working with postdoctoral fellows Dinis Pedro Calado, Gunther Galler, and others, created two strains of mice, a knockout lacking miR-150 and a transgenic mouse with miR-150 turned on early in development. The effects were strikingly complementary. The knockout displayed unusually high levels of a particular type of B cell, B1; the transgenic displayed almost no B1 cells, and B cell development in general was disrupted.
MiR-150 was clearly acting on the B cells, but it was not clear how—through what target. As it turned out, the researchers did not have to wait long for an answer. In the 1990s, Timothy Bender of the University of Virginia, working on sabbatical in Rajewsky’s lab, had generated a mouse lacking a transcription factor, c-Myb, that looked very similar to the miR-150 transgenic mice—abnormal B cell development and wholesale loss of B1 cells. And it was the mirror opposite of the knockout.
Meanwhile, using a bioinformatics approach, Rajewsky’s son, Nikolaus Rajewsky, then at New York University, had been scouring mRNAs to see which contained miR-150 binding sites. A noncoding region of the c-Myb mRNA had three such sites.
The researchers conducted a series of experiments to see if c-Myb was really the miR-150 target. They attached the noncoding region of the c-Myb gene to a luciferase reporter and introduced it into cultured cells containing or lacking miR-150. The reporter gene lit up only in those cells lacking miR-150. Further in vitro experiments showed that in the presence of miR-150, B cells underwent apoptosis.
Going back to the knockout mouse, Xiao and his colleagues found that c-Myb levels were higher than usual, as were amounts of the prosurvival factor BCL2, a downstream target of c-Myb. Antibodies specifically produced by B1 cells were also elevated. The transgenic mice showed other characteristics of interest. They had been engineered to produce different levels of miR-150. Xiao and colleagues found that the more miR-150 present, the lower the c-Myb concentration in the B cells and the fewer B cells produced by the mice.
Intriguingly, some of the mice exhibited the same reduction in c-Myb as found in mice with only one copy of the c-Myb gene. The heterozygotes are known to exhibit less than a 30 percent reduction in c-Myb, but it was not clear what effect such a relatively small reduction had on their phenotype. The researchers went back and looked at B cell development in the animals. Like the transgenic, the mice displayed an early and marked reduction in numbers of B cells produced in the bone marrow.
“This is perhaps the only paper so far where the target of a microRNA is functionally defined by actually changing the concentration of the target and then showing that you get the same phenotype as you get by operating through the microRNA,” Rajewsky said. “Until now, people only speculated about this.”